



How To Prevent Oxidative Damage In Your Mitochondria
tl;dr: Replacing linoleic acid with oleic acid in mitochondria protects against oxidative damage, resultant cell death, and the release of carcinogenic agents into the body (Zhong and Yin, 2015).

Background
Mitochondrial dysfunction appears to be the root cause of the metabolic syndrome, as I stated in the title to this post:
"The Cause of Metabolic Syndrome: Excess Omega-6 Fats (Linoleic Acid) in Your Mitochondria"
I noted that cardiolipin (CL) is the part of the mitochondria that is most subject to dysfunction, through oxidation of its linoleic acid (LA) constituents (it often contains four, on an industrial diet, and is known as tetralinoyl CL (TLCL)). I found a study (Zhong et al., 2014) that looked at cardiolipin that didn't contain four LA:
"The structure of L3OCL has one of the LA side chains replaced by an oleic acid [OA]. According to our chemical mechanism of 4-HNE formation from CL, cross-chain peroxyl radical reaction occurs between two adjacent side chains; thus the presence of an unreactive oleic acid side chain in L3OCL may disrupt this reaction and lead to less lipid electrophile production through this mechanism."
And sure enough, such cardiolipins produced less oxidatively-damaged products:
"As shown in Fig. 4, our data showed that the formation of similar electrophiles EAA-CL from L3OCL was indeed significantly suppressed compared to those from L4CL, highlighting the importance of this cross-chain reaction during the formation of reactive electrophiles."
Then I found a study (Lefkowith et al., 1985) that looked at mice that are deficient in LA. They replace the LA in their CL (which is only obtained through the diet) with OA (the main fat in olive oil. This is an omega-9 fat.) I summarized:
"In the next study we find that, sure enough, if you feed mice an essential fatty acid (EFA) "deficient" diet (0.05% LA) their cardiolipin used OA (which the body can produce) in place of LA. They shifted from 58% LA to 70% OA. The diet they were fed was also deficient in DHA, apparently."
Nevertheless:
"Animals on the partially EFA-deficient diet (restriction of linoleic acid to 10% of the control diet) were clinically normal, although hair loss did occur in some animals after 6 months." (Lefkowith et al., 1985)
A bit of DHA in the diet likely would have cured the hair loss.
In making my argument, I noted that:
"Now the hole in this argument, at the moment, is that I can't find a study that looks at OXLAM production and cell apoptosis in a LA-restricted diet."
Protecting Cardiolipin
Today I found this (Tyurina et al., 2012):
"Mitochondria targeting of non-peroxidizable triphenylphosphonium conjugated oleic acid protects mouse embryonic cells against apoptosis: Role of cardiolipin remodeling"
That's a bit of a mouthful, but what they did was pretty simple: they took OA (TPP- C18:1 in this study) and added it to CL, replacing the polyunsaturated fats (mostly LA) that are normally found there. OA acid is a monounsaturated fat, and is more resistant to oxidation—which is why olive oil is a good cooking fat. They did this in mouse embryonic cells (MEC), not in whole mice, as the next step was to give the cells a poison (actinomycin D, AcD) that causes CL oxidation and then apoptosis:
"Indeed, treatment with TPP- C18:1 markedly increased resistance of MEC to AcD induced apoptosis..."
Then, just to make sure, they introduced a substance (triacsin C) that blocked integration of OA into the cells:
"We found that triacsin C treated cells exhibited high sensitivity to AcD-induced apoptosis and abolished the anti-apoptotic effect of TPP-C18:1 "
Not, apparently, being familiar with the 1985 study mentioned above, they noted that replacing LA with OA didn't seem to have any impact on the function of the mitochondria:
"It is possible that enrichment of mitochondria with oleoyl-containing CL molecular species may affect interactions of this essential anionic phospholipid with a number of mitochondrial proteins such as cyt c oxidase [47,48], creatine kinase [49,50], ATP synthase [51,52] mitochondrial ADP carrier [53] resulting in altered functional status. Our previous work, however, demonstrated that depletion of 55% of endogenous CL in HeLa cells did not affect their growth rate, mitochondria biomarker proteins and levels of ATP and mitochondrial membrane potential [28]. Further, cyt c binding constant for tetra-oleoyl-CL was similar to that of tetra-linoleoyl-CL [54]."
So everything looked good. Not surprising, given the 1985 results.
However, that last sentence in that quote provides an additional, important clue. From Wikipedia:
"Cytochrome c is a highly conserved protein across the spectrum of species, found in plants, animals, and many unicellular organisms."
"Highly conserved" is a biological way of saying extremely important to the function of the electron transport chain (ETC), which means to life itself. This review of mitochondrial function—a neat read! (Stowe and Camara, 2009)—explains:
"Cardiolipin is a phospholipid found only in the IMM [inner-mitochondrial membrane] that anchors the mobile electron carrier cytochrome c to the IMM and optimizes the activity of electron-transport complexes, especially complex IV."
So a happy interaction between cyt. c and cardiolipin is extremely important, and if it breaks down, as mentioned in the previous post, the mitochondria breaks, perhaps killing the cell it's in (apoptosis, again).
Now the usual explanation for LA-containing cardiolipin is that since it's right next to mitochondrial complex I, which is where most ROS are created, it's constantly getting oxidized. This is a pretty good model, but it turns out there's another mechanism for ROS generation in the mitochondria.
"Cytochrome c-promoted cardiolipin oxidation generates singlet molecular oxygen." (Miyamoto et al., 2012)
Singlet oxygen is another type of ROS:
"Singlet oxygen is highly reactive, especially with organic compounds that contain double bonds."
Of course “organic compounds that contain double bonds” are PUFA like LA. Although, according to Wikipedia, singlet oxidation of PUFA is mostly an issue in chloroplasts, which are the mitochondria-analogue in plants.*
Anyway, it turns out that when cyt. c touches (which it does constantly!) CL with four LA, it can cause ROS release.
"In the present study, we have investigated whether cyt c–CL interaction could induce the generation of singlet oxygen. Chemical trapping and luminescence measurements at 1270 nm clearly showed the generation of singlet oxygen in cyt c incubation with CL liposomes. Quantitative analysis revealed the existence of at least two sources of singlet oxygen in our model, one directly related to CL hydroperoxide decomposition and another to CL oxidation." (Miyamoto et al., 2012)
And in their tests, this continued until there was no CL left!
"...Time dependent formation of singlet oxygen generation was followed by a parallel consumption of TLCL, reaching a saturation level when CL was totally consumed (Fig. 1C and 5B). The decrease in CL content was accompanied by the formation of CL monohydroperoxides especially at the first few minutes of the incubation, indicating that CL is being oxidized. This oxidation was probably activated by small amounts of CL hydroperoxides present in liposome preparations. It is important to note that Kagan’s group reported an enhancement of up to 1000-fold in the peroxidase activity of cyt c–CL complexes in the presence of fatty acid hydroperoxides, a process analogous to what happens with COX-1 and 2.[40] Thus similarly to fatty acid hydroperoxides, CL hydroperoxides can be used as a substrate necessary to activate cyt c peroxidase activity." (Miyamoto et al., 2012)
It's a standard observation that older mitochondria have fewer, oxidized, or missing CL. Adding CL or LA to mitochondria made dysfunctional by CL oxidation actually improves function, as it's better than none. It seems that what they're describing here is a mitochondrial death spiral...
[ P.S. "These reactive oxygen species readily attack the polyunsaturated fatty acids of the fatty acid membrane, initiating a self-propagating chain reaction." (Mylonas and Kouretas, 1999)]
LA-rich Cardiolipin are Uniquely Harmful
Now what’s interesting is that this is not a problem for all cardiolipin. Only for those comprised of specific fats.
"Moreover, singlet oxygen formation was specifically observed for tetralinoleoyl CL species and was not observed with monounsaturated and saturated CL species.” (Miyamoto et al., 2012)
This combination of LA-rich cardiolipin and cyt. c provides an alternative and unregulated source of ROS.
“Our results show that there are at least two mechanisms leading to singlet oxygen formation: one with fast kinetics involving the generation of singlet oxygen directly from CL hydroperoxide decomposition and the other involving CL oxidation. The contribution of the first mechanism was clearly evidenced by the detection of labeled singlet oxygen [18O2( 1 Δg)] from liposomes supplemented with 18-oxygen-labeled CL hydroperoxides. However quantitative analysis showed that singlet oxygen yield from CL hydroperoxides was minor ( < 5%) and that most of the singlet oxygen is formed from the second mechanism. Based on these data and previous findings we propose a mechanism of singlet oxygen generation through reactions involving peroxyl radicals (Russell mechanism) and excited triplet carbonyl intermediates (energy transfer mechanism).
"...This specificity is possibly related to the ability of linoleoyl [LA] acyl chains to induce structural rearrangements required for maximal activation of cyt c peroxidase activity [43] and also to the higher oxidizability of this fatty acid compared to oleoyl [OA] acyl chains." (Miyamoto et al., 2012)
That's pretty important. LA inclusion in CL causes ROS release, causes the release of toxic LA metabolites, and causes CL oxidation. Those LA metabolites induce oxidative damage potential "about three orders of magnitude higher in the presence of free fatty acid hydroperoxides ... than with hydrogen peroxide...." And it doesn't happen with CL that don't contain LA.
They sum:
"This process induces the formation of peroxyl radicals as well as excited triplet states, which are all known well established sources of singlet oxygen. The biological significance of singlet oxygen generated by the cyt c–CL complex needs to be further investigated. Nonetheless, our findings point to a potential role of singlet oxygen as an additional oxidant possibly involved in the pathway leading to cyt c release from mitochondria and induction of cell death." (Miyamoto et al., 2012)
Get your heart-healthy vegetable oils! (AHA 2016)
P.P.S.
It occurred to me that the demonstration of 4-HNE production (which is important for later posts) isn't demonstrated as strongly as I'd like.
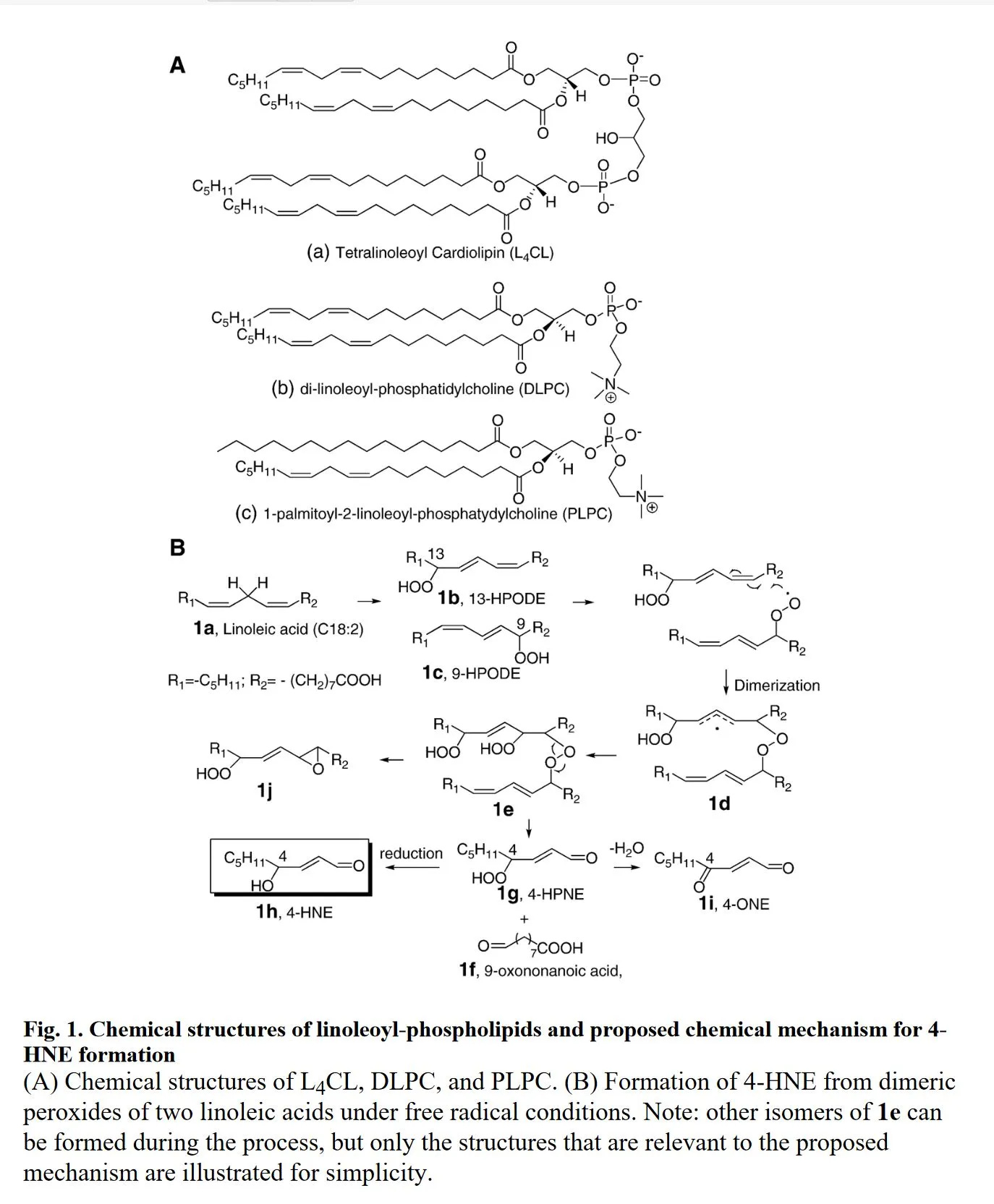
So here:
"...This novel mechanism for 4-HNE formation from CL oxidation in mitochondria may potentially have some physiological and pathophysiological significance. Generation of 4-HNE in vivo has been implicated in cardiovascular diseases, such as atherosclerosis [55], and neurodegenerative diseases including Alzheimer’s disease and Parkinson’s disease [5, 56–60]. L4CL [TLCL] is the major CL species in the mitochondria of most mammalian tissues and it constitutes more than 70% in the heart [61]. Thus formation of 4-HNE via this novel mechanisms is likely important. 4-HNE alters multiple essential functions of brain mitochondria, which plays a pivotal role in initiation and progression of neurodegenerative diseases [62]. 4-HNE also induces mitochondrial dysfunction and aberrant axonal outgrowth in adult sensory neurons that mimics features of diabetic neuropathy [63]....
"...In summary, we have provided evidence that 4-HNE and related reactive lipid electrophiles can be generated through a novel chemical mechanism of cardiolipin oxidation under free radical conditions in vitro and in vivo. This mechanism links 4-HNE generation to cardiolipin oxidation in mitochondria where abundant ROS are generated." (Liu et al., 2011)
*LA is of course a PUFA that comes exclusively from plants, as animals don't produce it. I'll try not to read to much into that, but it turns out that LA in plants is subject to oxidative damage from singlet oxygen:
"Singlet Oxygen Is the Major Reactive Oxygen Species Involved in Photooxidative Damage to Plants" (Triantaphylidès et al., 2008)
The notion that we may have imported this problem from plants is an interesting one, but definitely an aside!
Original post February 23, 2016.
References
American Heart Association. (2016, March 1). Healthy Cooking Oils 101 [Advertisement]. American Heart Association. https://web.archive.org/web/20160301070355/http://www.heart.org/HEARTORG/HealthyLiving/HealthyEating/SimpleCookingwithHeart/Healthy-Cooking-Oils-101_UCM_445179_Article.jsp#.VtU-4nbP1qY
Lefkowith, J. B., Flippo, V., Sprecher, H., & Needleman, P. (1985). Paradoxical Conservation of Cardiac and Renal Arachidonate Content in Essential Fatty Acid Deficiency. Journal of Biological Chemistry, 260(29), 15736–15744.
Miyamoto, S., Nantes, I. L., Faria, P. A., Cunha, D., Ronsein, G. E., Medeiros, M. H. G., & Di Mascio, P. (2012). Cytochrome C-Promoted Cardiolipin Oxidation Generates Singlet Molecular Oxygen. Photochemical & Photobiological Sciences, 11(10), 1536. https://doi.org/10.1039/c2pp25119a
Mylonas, C., & Kouretas, D. (1999). Lipid Peroxidation and Tissue Damage. In Vivo (Athens, Greece), 13(3), 295–309.
Stowe, D. F., & Camara, A. K. S. (2009). Mitochondrial Reactive Oxygen Species Production in Excitable Cells: Modulators of Mitochondrial and Cell Function. Antioxidants & Redox Signaling, 11(6), 1373–1414. https://doi.org/10.1089/ars.2008.2331
Triantaphylidès, C., Krischke, M., Hoeberichts, F. A., Ksas, B., Gresser, G., Havaux, M., Van Breusegem, F., & Mueller, M. J. (2008). Singlet Oxygen Is the Major Reactive Oxygen Species Involved in Photooxidative Damage to Plants. Plant Physiology, 148(2), 960–968. https://doi.org/10.1104/pp.108.125690
Tyurina, Y. Y., Tungekar, M. A., Jung, M.-Y., Tyurin, V. A., Greenberger, J. S., Stoyanovsky, D. A., & Kagan, V. E. (2012). Mitochondria Targeting of Non-Peroxidizable Triphenylphosphonium Conjugated Oleic Acid Protects Mouse Embryonic Cells Against Apoptosis: Role of Cardiolipin Remodeling. Febs Letters, 586(3), 235–241. https://doi.org/10.1016/j.febslet.2011.12.016
Zhong, H., Lu, J., Xia, L., Zhu, M., & Yin, H. (2014). Formation of Electrophilic Oxidation Products from Mitochondrial Cardiolipin in Vitro and in Vivo in the Context of Apoptosis and Atherosclerosis. Redox Biology, 2, 878–883. https://doi.org/10.1016/j.redox.2014.04.003
Zhong, H., & Yin, H. (2015). Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in cancer: Focusing on mitochondria. Redox Biology, 4, 193–199. https://doi.org/10.1016/j.redox.2014.12.011
Thank you for this - what of it I can follow is illuminating. It leaves me wondering about OA, though. Just yesterday I listened to the podcast with Brad Marshall and Paul Saladino on why olive oil is worse than, say, butter or beef tallow, and OA is a culprit there in other ways. It's obviously miles better than LA, but what should we conclude?
How does one get PUFA-free olive oil ? It does contain too much linoleic acid after all